
A közlemény célja, hogy összefoglalja a vascularis simaizomsejtek kálíum-ioncsatornáinak biofizikai és farmakológiai tulajdonságait. Az érsimaizom sejtek membránpotenciálját jelentősen meghatározza a sejtmembrán K+-vezetőképessége. A membránpotenciál kialakításában több K+ csatornatípus játszik szerepet. A feszültségfüggő K-csatornák a nyugalmi membránpotenciál kialakításában/helyreállításában vesznek részt. A kalciumfüggő K+-csatornákat az intracelluláris Ca2+-szint szabályozza és a simaizomtónus beállításában tulajdonítanak nekik szerepel. Közülük a kis konduktanciájú K+-csatornák az endothelsejtek felől érkező hyperpolarizáló hatások iránt érzékenyek. Az ATP-függő K+-csatornák aktivációja az izomsejt metabolikus állapotának függvénye. Az extracelluláris K+ - szint változásait külön K+-esatornatípus érzékeli. A fenti ioncsatornák közül az utóbbi időben megkülönböztetett figyelem irányult az ATP-szenzitív és a Ca2+- függő K+-ioncsatornákra. Fiziológiás körülmények között a vascularis simaizom kálium csatornáinak összehangolt működése jelenti a perifériás vérnyomás- és a regionális véráramlás-szabályozás egyik legfontosabb elemét. Mivel a K+-csatornák számos vazoaktív anyag támadáspontját képezik, ezen csatornák működésének ismerete segíti a hatásmechanizmusok megértését. A közleményben ismertetésre kerübiek a szerző és munkatársai újszülött malacok agyi arterioláin végzett kísérletei, melyek során megfigyelték az anoxia/reperfúzió szelektív K+-csatorna károsító hatását.
A simaizom K+-csatornái funkcionális károsodásának ismerete lehetővé teszi egyes pathomechanizmusok tisztázását, és támpontot szolgáltat új terápiás beavatkozások megtervezéséhez.
Érbetegségek: 1998/1. 1-5. oldal
KULCSSZAVAK
vascularis simaizom, K+- csatorna, arteriola, ischaemia
A vascularis simaizom tónusát alapvetően a myoplasma szabad Ca2+-koncentrációja határozza meg. A sejten belüli Ca2+-szint alakulása számos mechanizmus eredménye. Közülük meghatározó jelentőséget tulajdonítanak azoknak a hatásoknak, amelyek a sejtmembrán ioncsatornáinak vezetőképességét szabályozzák. A Ca2+ által átjárható csatornák ionáramai közvetlenül hatnak a myogén tónusra, míg azt az egyéb töltött részecskék áramlása áttételeken keresztül befolyásolja.
Az utóbbi évtizedben megkülönböztetett figyelem irányult az értónus szabályozása szempontjából (is) nagy jelentőségű K+-áramok és K+-csatornák vizsgálatára. A sejtmembrán K+-áramai - melyek a membrán K+-vezetőképességétől függenek - döntő mértékben meghatározzák az intra- és extracelluláris tér között mérhető membránpotenciál értékét. A K+-vezetőképesség szabályozása szempontjából különösen fontosak a K+-ra szelektív ioncsatornák. Ezen csatornák vezetőképessége nyitvatartási valószínűségük (aktivált/deaktivált állapotuk időbeni aránya) függvénye. A K+- vezetőképesség és ez által a membránpotenciál változása közvetett hatást gyakorol a sejten belüli Ca2+-koncentrációra, amely közvetlenül szabályozza a myogén tónust.
Az extra- és intracelluláris térben található ionok mozgásához a legnagyobb hajtóerőt az eltérő koncentráció, valamint a nyugalmi membrán potenciál (és az ezzel arányos elektromos térerő) jelenti. A simaizomsejtekben az intracelluláris K+-koncentráció az extracelluláris koncentrációnak kb. 25-szöröse, A nyugalmi membránpotenciál -45-60 mV (ami pozitívabb a K+ Nernst-egyenletből számítható egyensúlyi potenciáljánál). így nyugalomban a K+-ra sejtből kifelé mutató mozgatóerő hat. A sarcolemma K+-csatornáinak megnyílása kifelé irányuló pozitív töltésáramlást és hyperpolarizációt eredményez. Amennyiben a membrán permea- bilitása szelektíven, csak a K+-ionokra nő meg, és a többi ionra vonatkozó vezetőképesség változatlan marad, a membrán potenciál a K+-ionok egyensúlyi potenciáljához (-90 mV) közeli értéken stabilizálódik.
A membrándepolarizáció serkenti, míg a hyperpolarizáció gátolja a feszültségfüggő (T- és L-típusú) Ca2+-csatornák aktivitását, és jelentősen meghatározza az intracelluláris szabad Ca2+-szint alakulását. A K+-csatornák aktiválása során membrán depolarizáció jön létre és egyidejűleg további intracelluláris mechanizmusok aktiválódnak. Gátlás alá kerül az inositol trifoszfát (IP3) képződés, ennek következtében kevesebb Ca2+ mobilizálódik az intracelluláris raktárakból. Csökken a Ca2+ kontraktilis fehérjékre gyakorolt hatása. A hyperpolarizáció serkenti a transzmembrán Na+/Ca2+ cserét, és ezzel csökken az intracelluláris szabad Ca2+-szint. Ezen szerteágazó hatások következményeként a vascularis simaizomsejt vagy relaxált állapotba kerül, vagy csökken a kontrakciót serkentő tényezők iránti érzékenysége.
A K+-csatornák a membrán csatornák heterogén csoportját képezik. Az ismert K+-csatornák strukturális jellemzése, aminosav szekvenciák azonosítása nagy ütemben halad. Struktúrájuk alapján a K+-csatornák három nagy csoportba sorolhatók. Az azonos csoportokba tartozó csatornafehérjék 6, 2, illetve 1 transzmembrán szegmenssel rendelkeznek. A struktúra szerinti osztályozásnál elterjedtebb azonban a K+- csatornák funkció szerinti besorolása.
Mind a kálium-ioncsatornák blokkolása, mind azok aktiválása alapvetően megváltoztatja a simaizom ingerlékenységét. A K+-csatornákat aktiváló szerek simaizom hyperpolarizációt és vazodilatációt okozó hatása különböző szervekből nyert artéria és véna preparátumokon végzett kísérletekből hosszú idő óta ismert. Az is bizonyított, hogy a K+-csatornák megnyitásával ellensúlyozható az egyébként depolarizációt okozó szerek hatása is. (Együttes alkalmazásuk a membrán repolarizációjához vezet.)
Intakt artériákon és vénákon végzett farmakológiai és izolált simaizom sejteken végzett patch-clamp kísérletek alapján (az utóbb említett eljárás lehetőséget teremt az egyes ioncsatornák szelektív és "individuális" vizsgálatára) az érsimaizom sejteken 4 különböző káliumcsatorna típust azonosítottak (melyek további alcsoportokba sorolhatók). A négy nagy csoport jellemző tulajdonságai a következők (bővebben: 4, 13, 14, 15):
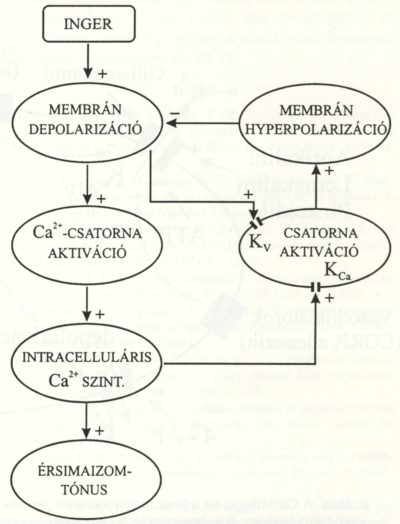
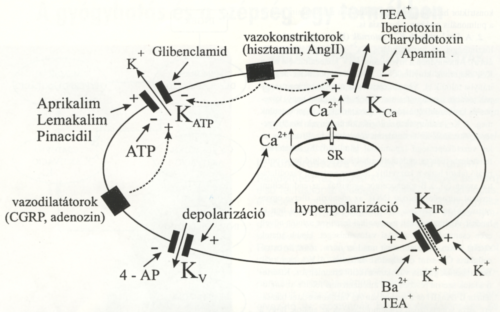
1. ábra.
A vascularis simaizomsejt membránjában található K+-csatornák és a legfontosabb aktiváló mechanizmusaik, valamint gátlásuk lehetőségei. Az ábrára vonatkozó információt a szövegben az egyes csatorna típusok leírása tartalmazza.
2. ábra.
A Ca2+-függő és a feszültség vezérelt (Kca és Kv) K+-csatornák szerepe az érsimaizom tónusának szabályozásában. A sejtmembrán K+ vezetőképességének változása elsősorban a membránpotenciálon keresztül szabályozza az intracelluláris Ca2+ - szintet. A depolarizáció hatására a Kv csatornák, míg a Ca2+ szint emelkedés hatására a Kca csatornák aktiválódnak, és stabilizálódik a membránpotenciál.
1. A feszültségfüggő kálium (Kv) csatornákat a membrán depolarizáció aktiválja. Funkciójuk valószínűleg a nyugalmi membránpotenciál visszaállítása, illetve a depoiarizáló hatások ellensúlyozása (a membránpotenciál stabilizálása) (1. és 2. ábra). Ezek az ioncsatornák elektrofiziológiai módszerekkel, vezetőképességük és az aktivizálás dinamikája alapján még további altípusokba sorolhatók. A Kv csatornák aktivitása hatásosan és szelektíven gátolható kis koncentrációban (0,2-1,0 mmol/L) alkalmazott 4-aminopiridinnel (4- AP). Valószínűleg Kv csatornák zárásával fejti ki vazokonstriktor hatását egyes érszakaszokon a hisztamin, illetve a pulmonális erekben a hypoxia is.
2. A Ca2+-aktivált K+-csatornák (Kca) nyitvatartási valószínűségét az intracelluláris Ca2+-szint szabályozza (1. ábra). Amennyiben a myoplasma Ca2+ koncentrációja nő, a Kca csatornák kinyílnak, és a membránpotenciál negatív irányba tolódik el. Vezetőképességük alapján ezek a csatornák további osztályokba sorolhatók. Ez alapján nagy, közepes és kis vezetőképességű Kca csatornákat különböztetünk meg. A Kca csatornák fiziológiás körülmények között szerepet játszanak az érsimaizom tónusának beállításában. A kis vezetőképességű (apaminnal gátolható) Kca-csatornák feltételezhetően az endotheliumból felszabaduló hyperpolarizáló faktor hatását közvetítik. Számos endogén vazodilatátor anyag (pl. a P-adrenoceptor agonisták, az endothelium eredetű nitrogénmonoxid [NO], az adenozin, illetve egyes zsírsavak) is részben a Kca csatornák aktiválásával fejti ki hatását. A simaizomsejt mechanikai feszülése szintén növeli ezen csatornák nyitvatartási valószínűségét. Újabb adatok szerint a Kca-ioncsatornákat mind az intra-, mind az extra- celluláris CO-szint emelkedése aktiválja. A Kca csatornák aktivitásának gátlása vazokonstrikciót eredményez. Kísérleti adatok szerint részben ez a mechanizmus felelős az angiotenzin II (AngII) és a tromboxán A2 vazokonstriktor hatásáért. Arra vonatkozóan ellentmondásosak az adatok, hogy a nyitott Kca-ioncsatornák fiziológiás körülmények között részt vesznek-e a nyugalmi vascularis tónus fenntartásában.
A Kca-csatornák funkcionális vizsgálatát sokáig nehezítette, hogy nem állt rendelkezésre ezen csatornáknak sem szelektív exogén agonistája, sem antagonistája. Az utóbbi időben két skorpió toxin, a charybdotoxin és az iberiotoxin hatásos Kca antagonistának bizonyult. Segítségükkel sikerült tisztázni, hogy bizonyos érszakaszokon a Kca-csatornák aktivált állapota is felelős az érsimaizomsejtek nyugalmi tónusának fenntartásáért.
3. Az ATP-szenzitív K+ (KATP) csatornák elnevezésében tükröződik az a megfigyelés, hogy ezek a csatornák az intracelluláris ATP szint csökkenésére, illetve az ATP/ADP arány változására érzékenyek. Az is bizonyított, hogy ezeket a csatornákat más vazoaktív anyagok, például peptidek, szöveti metabolitok és farmakonok is aktiválják (1. ábra). A KATP-csatornáknak számos endogén ligandja ismert. Részben, vagy teljes egészében ezen csatornák megnyitásával lejti ki vazodilatátor hatását többek között a calcitonin-gén relációs peptid (CGRP), a prosztaciklinek, az adenozin és a szomatosztatin. A CGRP elsősorban az érző idegvégződésekből szabadul fel, és egyike az ismert leghatásosabb vazodilatátor anyagoknak.
A szulfonilurea származékokról ismert, hogy gátolják a KATP-csatornákat. Közülük a leggyakrabban használt KATP antagonista a glibenclamid, amely |xmol-os koncentrációban hatásosan gátolja mind az exogén (aprikalim, lemacalim, cromakalim, vagy pinacidil), mind az endogén (pl. CGRP,adenozin) agonisták hatását. Elsősorban coronaria artériákból nyert simaizomsejteken végzett elektrofiziológiai vizsgálatokból tudjuk, hogy a vazopresszin, angiotenzin és az endothelin-1 gátolja a KATP-csatornák ionáramát. Feltételezhetően ez is magyarázza az említett anyagok vazokonstriktor hatását.
Fiziológiás körülmények között több árterületen a simaizomsejtekben az ATP szint elegendően magas ahhoz, hogy a KATP csatornák zártak maradjanak, azaz a nyugalmi membránpotenciál kialakításában ezeknek a csatornáknak elhanyagolható a szerepük. Ezzel megegyezik mindazoknak az in vitro és in vivo kísérleteknek az eredménye, amelyekben igazolták, hogy a KATP-csatornák gátlása fiziológiás körülmények között nincs hatással sem az agyi erek simaizomsejtjeinek nyugalmi membránpotenciáljára, sem az agyi arteriolák tónusára. Ezzel szemben, a coronaria ereken glibenclamid kezelés hatására vazokonstrikció alakul ki, jelezvén, hogy a bazális értónus fenntartásában szerepet játszanak a nyitott KATP-csatornák is. A KATP-csatornáknak azokon az árterületeken tulajdonítanak különösen nagy szerepet a lokális vérellátás szabályozásában, ahol a szöveti perfúzió szorosan követi a metabolikus igényeket. így a központi idegrendszerben, a szív- és vázizomzatban a KATP-csatornák aktiválása valószínűleg fiziológiás körülmények között is részt vesz a vazodilatációs mechanizmusokban. Egyrészt úgy, hogy a megnövekedett metabolikus aktivitás hatására az erek közvetlen környezetében megnő az adenozin, a K+ és a proton koncentráció. Közülük az adenozin aktiválja KATP-csatornákat. Másfelől a megemelkedett 02-felhasználás eredményeként magában az érsimaizomsejtekben is csökken az ATP termelés, aminek következtében megnyílhatnak a sejt KATP-csatornái. Ezek a hatások összegződhetnek, ami további KATP-csatorna aktivációt eredményezhet.
4. Az inward rectifier K+ -csatornák (Kir) elnevezésében az tükröződik, hogy rajtuk keresztül a K+ elsősorban a sejt belseje felé áramlik (1. ábra). Ezeket a csatornákat a membrán hyperpolarizációja aktiválja. A Kir szerepe a membránpotenciál stabilizálásában jelentős. A Kir-csatornák az extra- celluláris K+ koncentráció változásaira is érzékenyek. Aktivitásuk bárium ionokkal (Ba2+) és tetraetilammóniummal (TEA) gátolható. Az intersticium változó kálium koncentrációját hosszú idő óta úgy tekintik, mint amely szerepet játszik a lokális véráramlás és a metabolikus igények összekapcsolásában. A pia matererei a lokális K+ koncentráció megemelkedésére (20 mmol/L-es határig) vazodilatációval válaszolnak. Ennek fiziológiás körülmények között is lehet szabályozó szerepe, hiszen az aktivált idegsejtek iC-ot veszítenek és környezetükben emelkedik a K+ koncentráció.
A cardiovascularis megbetegedések pathomechanizmusának vizsgálatában az utóbbi években kétségtelenül az endotheliális diszfunkciók jelentették a központi kérdést. Ezzel párhuzamosan azonban kiderült, hogy egyes kórképekben a simaizomsejtek ioncsatornái is strukturális és funkcionális változásokon mennek keresztül. A K+-csatornák szisztematikus vizsgálata, tulajdonságaik pontos jellemzése az utóbbi évtizedben kezdődött meg. A vascularis simaizom K+-ion- csatornáira vonatkozó ismereteket az elmúlt időszakban számos részletes tanulmány összegezte (4, 6, 13-15). Az összefoglalókból egyre részletesebben kirajzolódik, hogy fiziológiás körülmények között mind az artériák (15), mind a vénák (14) vazomotor funkcióiban döntő szerepük van a K+-ioncsatornák működésének (4).
Ezeknek az ioncsatornáknak az egyes kórállapotokhoz köthető elváltozásairól azonban még igen hiányosak az ismeretek (5, 8, 16). A továbbiakban röviden áttekintjük a vascularis simaizom K+-csatornáinak egyes, kísérletesen létrehozott kórképekhez köthető elváltozásait. Saját, újszülött malacok agyi arterioiáin végzett, in vivo kísérletek alapján bemutatjuk, hogy miként befolyásolja az anoxia/reperfúzió a K+-csatornák működését.
Elsősorban állatkísérletes modellekből ismerjük, hogy egyes kórképekben változik a Kca-csatornák érzékenysége, így például hypercholesterinaemiában a Kca-csatornák érzékenysége megnő, potencírozódik az agonisták hatása. Hasonló változást figyeltek meg só-diétával kialakított hypertenziós patkányok artériáin is (9, 17). Az agyi rezisztencia erek KATP-csatornáinak aktivitása kísérletesen létrehozott diabetes mellitusban csökken (12). A KATP-csatornákat aktiváló szerek hatástalanok maradnak, míg nitrovazodilatátorokkal továbbra is értágulat váltható ki.
A krónikus megbetegedések mellett egyéb hatások is módosítják a KATP-csatornák funkcióját. Újszülött malacok agyi arterioláin végzett kísérleteink során megvizsgáltuk, hogy globális agyi ischaemia és reperfúzió miként befolyásolja a KATP-csatornák ingerelhetőségét (3). Az ioncsatornák direkt aktiválásához aprikalimot, míg indirekt aktiválásukhoz CGRP-t és iloprostot (amely stabil porstaklin analóg) használtunk (1, 3, 11). A kiindulási körülmények között ezek a szerek kb. 100 µm nyugalmi átmérőjű arteriolákon dózis függő vazodilatációt okoztak. Hatásukat glibenclamiddal minden esetben fel lehetett függeszteni. Tíz perces teljes agyi ischaemiát és reperfúziót követően 1-4 órán keresztül vizsgáltuk a KATP-csatornák aktiválhatóságát. Eredményeink szerint az anoxiát/reperfúziót követő átmeneti hypo-, majd hyperperfúzió után 30-40 perccel az arteriolák átmérője már nem különbözött a kiindulási értéktől. Ezzel szemben az agyi arteriolák KATP-csatorna aktiválók iránti érzékenysége nagy mértékben csökkent. Egyidejűleg azonban változatlan maradt más vazodilatátor anyagok, például nitroprusszid nátrium, illetve prosztaglandin E2 iránti dilatációs készségük.
A KATP csatornák funkcionális károsodásának mechanizmusát csak részben sikerült tisztáznunk. Farmakológiai eszközökkel végzett vizsgálatainkból arra következtettünk, hogy a megváltozott funkcióért - legalábbis részben - a reperfúzió során keletkezett szabadgyökök tehetők felelősség. Más árterületeken végzett vizsgálatokban kimutatták azt is, hogy a hypoxia önmagában is gátolja az ATP szenzitív K+-csatornák ingerelhetőségét.
Szintén újszülött malacok agyi arterioláin vizsgáltuk, miként hat az anoxia a Kca-csatorna funkcióra (2). A Kca-csatornák aktiválása eredményeként arteriola dilatáció alakult ki. Vizsgálataink azt mutatták, hogy a Kca-csatornák az anoxiás stressznek jobban ellenállnak, mint a Kca-csatornák, hiszen ingerelhetőségük 10 perces teljes ischaemia és reperfúzió után sem változott meg számottevően.
Újabb vizsgálatok szerint a reaktív szabadgyökök is részben a Kca aktiválásán keresztül hatnak, ami magyarázata lehet az anoxiát követő és a szabadgyökképződéssel összefüggő hyperaemiának.
A fenti megállapítások igazolják, hogy a vascularis simaizomsejtek K+-ioncsatornái fontos szerepet játszanak az ér- tónus és ezzel együtt a véráramlás szabályozásában. A csatornafunkciók ismerete, egyes kórképekhez köthető elváltozásainak jellemzése segíti a kóros állapotok megértését, és lehetőséget teremt új gyógyszerek tervezéséhez és terápiás eljárások bevezetéséhez (7, 10, 18).
Irodalom
- Bari F., Errico, R. A., Louis, T. M., Busija, D. W.: Interaction between ATP-sensitive K+ channels and nitric oxidé on piai arterioles in piglets. J. Cerebral Blood Flow Metab. 16: 1158-64. (1996)
- Bari F., Louis, T. M., Busija, D. W.: Calcium-activated K+ channels in cerebral arterioles in piglets are resistant to ischemia. .). Cerebral Blood Flow Metabol. (in press). (1997)
- Bari F., Louis. T. M., Meng, W., Busija. D. W.: Global ischemia impairs ATP-sensitive K+ channel function in cerebral arterioles in piglets. Stroke 27:1874-81. (1996)
- Braycen, J. E.: Potassium channels in vascular smooth muscle. Clin. Experiment Pharmacol. Physiol. 23:1069-76. (1996)
- Cameron. J. S., Baghdady, R.: Role of ATP sensitive potassium channels in long term adaptation to metabolic stress. Cardiovasc. Res. 28:788-96. (1994)
- Challinor-Rogers, .J. L. McPherson, G.A.: Potassium channel openers and other regulators of KATP channel. Clin. Experiment Pharmacol. Physiol. 21: 583-597. (1994)
- Escande. D.. Henry, P.: Potassium channels as pharmacological targets in cardiovascular medicine. Eur. Heart Journal. 14 Suppl. B: 2-9. (1993)
- Faraci, F. M.. Sobey. C. G.: Potassium channels and the cerebral circulation. Clin. Experiment Pharmacol. Physiol. 23:1091- 95. (1996)
- Kitanozo, T., Heistad, D. D.. Faraci, F. M.: ATP-sensitive potassium channels in the basilar artery during chronic hyperten- sion. Hypertension 22: 677-81. (1993)
- Lawson. K.: Potassium channel activation: apotential therape- utic approach? Pharmacol. Therapeutics. 70: 39-63. (1996)
- Ouis. T. M" Meng, W.. Bari F., Errico, R. A.. Busija. D. W.: Ischaemia reduces CGRP-induced cerebral vascular dilation in piglets. Stroke. 27: 134-139 (1996).
- Mayhan, W. G.. Faraci, F. M.: Responses of cerebral arterioles in diabetic rats to activation of ATP-sesitive potassium channels. Am. J. Physiol. 265: H152-H157. (1993)
- McPherson. G. A: Current trends in the study of potassium channel openers. General Pharmacol. 24(2):275-81.
- Monos E.. Bérezi V.. Nádasy Gy.: Local control of veins: bio- mechanical, metabolic and humorai aspeets. Physiol. Rev. 75:611-666. (1995)
- Nelson, M. T.. Quayle, .J. M.: Physiological roles and properti- es of potassium channels in arterial smooth muscle. Am. J. Physiol. 268:C799-822. (1995)
- Pierce G. N.. Czubryt, M. P.: The contribution of ionic imbalance to ischemia/reperfusion-induced injury. J. Mol. Cell. Cardiol. 27:53-63. (1995)
- Rusch, N. .J., Liu, Y., Pleyte, K. A.: Mechanisms for reguládon of arterial tone by Ca2+-dependent K+ channels in hypertension. Clin. Experiment Pharmacol. Physiol. 23: 1077-81. (1996)
- Zhang, H.. Cook, D.: Cerebral vascular smooth muscle potassium channels and their possible role in the management of vasosplasm. Pharmacol. Toxicol. 75:327-36. (1994)
Dr. Bari Ferenc
SZOTE Élettani Intézete
6720 Szeged, Dóm tér 10.
Érbetegségek: 1998/1. 1-5. oldal
